
- DNA Replication
- Active Transport
- Cellular Receptors
- Endocytosis and Exocytosis
- Enzyme Inhibition
- Enzyme Kinetics
- Protein Structure
- Transcription of DNA
- Translation of DNA
- Anaerobic Respiration
- Electron Transport Chain
- Gluconeogenesis
- Calcium Regulation
- External Balance of Potassium
- Internal Balance of Potassium
- Sodium Regulation
- Cell Membrane
- Endoplasmic Reticulum
- Golgi Apparatus
- Mitochondria
- Blood Vessels
- Cellular Adaptations
- Epithelial Cells
- Muscle Histology
- Structure of Glands
- Control of Stroke Volume
- Control of Heart Rate
- Cardiac Cycle
- Cardiac Pacemaker Cells
- Conduction System
- Contraction of Cardiac Muscle
- Ventricular Action Potentials
- Blood Flow in Vessels
- Control of Blood Pressure
- Capillary Exchange
- Flow In Peripheral Circulation
- Venous Return
- Cardiac Muscle
- Hepatic Circulation
- Skeletal Muscle
- Airway Resistance
- Lung Volumes
- Mechanics of Breathing
- Gas Exchange
- Oxygen Transport in The Blood
- Transport of Carbon Dioxide in the Blood
- Ventilation-Perfusion Matching
- Chemoreceptors
- Cough Reflex
- Neural Control of Ventilation
- Respiratory Regulation of Acid-Base Balance
- Responses of The Respiratory System to Stress
- Regulation of Saliva
- Secretion of Saliva
- Gastric Acid Production
- Gastric Mucus Production
- Digestion and Absorption
- Histology and Cellular Function of the Small Intestine
- Absorption in the Large Intestine
- Large Intestinal Motility
- Bilirubin Metabolism
- Carbohydrate Metabolism in the Liver
- Lipid Metabolism in the Liver
- Protein and Ammonia Metabolism in the Liver
- Storage Functions of the Liver
- Bile Production
- Function of The Spleen
- Exocrine Pancreas
- Somatostatin
- Proximal Convoluted Tubule
- Loop of Henle
- Distal Convoluted Tubule and Collecting Duct
- Storage Phase of Micturition
- Voiding Phase of Micturition
- Antidiuretic Hormone
- Renin-Angiotensin-Aldosterone System
- Urinary Regulation of Acid-Base Balance
- Water Filtration and Reabsorption
- Development of the Reproductive System
- Gametogenesis
- Gonadotropins and the Hypothalamic Pituitary Axis
- Menstrual Cycle
- Placental Development
- Fetal Circulation
- Maternal Adaptations in Pregnancy
- Cells of the Nervous System
- Central Nervous System
- Cerebrospinal Fluid
- Neurotransmitters
- Peripheral Nervous System
- Action Potential
- Excitatory and Inhibitory Synaptic Signalling
- Resting Membrane Potential
- Synaptic Plasticity
- Synaptic Transmission
- Ascending Tracts
- Auditory Pathway
- Consciousness and Sleep
- Modalities of Sensation
- Pain Pathways
- Sensory Acuity
- Visual Pathway
- Descending Tracts
- Lower Motor Neurones
- Muscle Stretch Reflex
- Upper Motor Neurones
- Aqueous Humour
- Ocular Accommodation
- Thyroid Gland
- Parathyroid Glands
- Adrenal Medulla
- Zona Glomerulosa
- Zona Fasciculata
- Zona Reticularis
- Endocrine Pancreas
- The Hypothalamus
- Anterior Pituitary
- Posterior Pituitary
- White Blood Cells – Summary
- Barriers to Infection
- Infection Recognition Molecules
- Phagocytosis
- The Complement System

Antigen Processing and Presentation
- Primary and Secondary Immune Responses
- T Cell Memory
- Acute Inflammation
- Autoimmunity
- Chronic Inflammation
- Hypersensitivity Reactions
- Immunodeficiency
- Types of Immunity
- Antibiotics
- Viral Infection
- Blood Groups
- Coagulation
- Erythropoiesis
- Iron Metabolism
- Mononuclear Phagocyte System
Original Author(s): Antonia Round Last updated: 17th July 2023 Revisions: 9
- 1 Antigen Presentation
- 2.1 MHC Class I Molecules
- 2.2 MCH Class II Molecules
- 3.1 T Cell Receptors
- 3.2 Co-Receptors
- 4 Clinical Relevance – Autoimmune disease
T cells can only recognise antigens when they are displayed on cell surfaces. This is carried out by Antigen-presenting cells (APCs) , the most important of which are dendritic cells, B cells, and macrophages. APCs can digest proteins they encounter and display peptide fragments from them on their surfaces for other immune cells to recognise.
This process of antigen presentation allows T cells to “see” what proteins are present in the body and to form an adaptive immune response against them. In this article, we shall discuss antigen processing, presentation, and recognition by T cells.
Antigen Presentation
Antigens are delivered to the surface of APCs by Major Histocompatibility Complex (MHC) molecules. Different MHC molecules can bind different peptides. The MHC is highly polygenic and polymorphic which equips us to recognise a vast array of different antigens we might encounter. There are different classes of MHC, which have different functions:
- MHC class I molecules are found on all nucleated cells (not just professional APCs) and typically present intracellular antigens such as viruses.
- MHC class II molecules are only found on APCs and typically present extracellular antigens such as bacteria.
This is logical because should a virus be inside a cell of any type, the immune system needs to be able to respond to it. This also explains why pathogens inside human red blood cells (which are non-nucleated) can be difficult for the immune system to find, such as in malaria.
Whilst this is the general rule, in cross-presentation extracellular antigens can be presented by MHC class I, and in autophagy intracellular antigens can be presented by MHC class II.
Antigen Processing
Before an antigen can be presented, it must first be processed . Processing transforms proteins into antigenic peptides.
MHC Class I Molecules
Intracellular peptides for MHC class I presentation are made by proteases and the proteasome in the cytosol, then transported into the endoplasmic reticulum via TAP (Transporter associated with Antigen Processing) to be further processed.
They are then assembled together with MHC I molecules and travel to the cell surface ready for presentation.
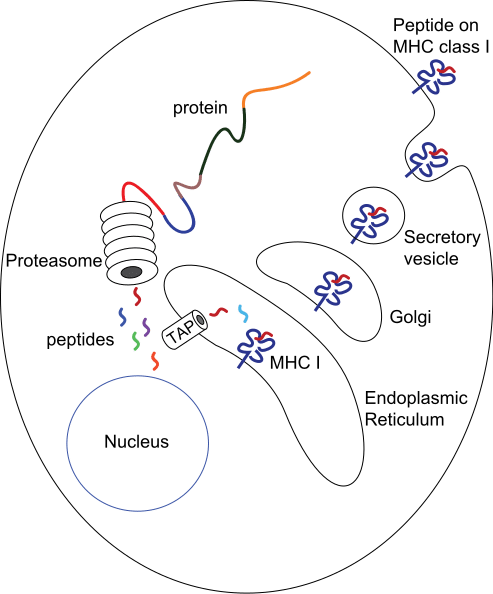
Fig 1 – Diagram demonstrating the production of peptides for MHC class I presentation
MCH Class II Molecules
The route of processing for exogenous antigens for MHC class II presentation begins with endocytosis of the antigen. Once inside the cell, they are encased within endosomes that acidify and activate proteases, to degrade the antigen.
MHC class II molecules are transported into endocytic vesicles where they bind peptide antigen and then travel to the cell surface.
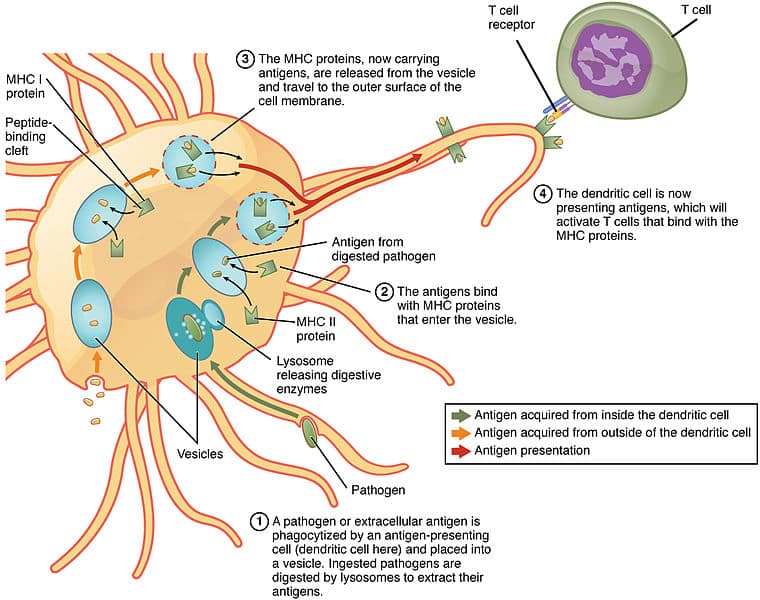
Fig 2 – Diagram showing processing of antigens for MHC Class II presentation by a dendritic cell
The antigen presented on MHCs is recognised by T cells using a T cell receptor (TCR) . These are antigen-specific .
T Cell Receptors
Each T cell has thousands of TCRs , each with a unique specificity that collectively allows our immune system to recognise a wide array of antigens.
This diversity in TCRs is achieved through a process called V(D)J recombination during development in the thymus. TCR chains have a variable region where gene segments are randomly rearranged, using the proteins RAG1 and RAG2 to initiate cleavage and non-homologous end joining to rejoin the chains.
The diversity of the TCRs can be further increased by inserting or deleting nucleotides at the junctions of gene segments; together forming the potential to create up to 10 15 unique TCRs.
TCRs are specific not only for a particular antigen but also for a specific MHC molecule. T cells will only recognise an antigen if a specific antigen with a specific MHC molecule is present: this phenomenon is called MHC restriction .
Co-Receptors
As well as the TCR, another T cell molecule is required for antigen recognition and is known as a co-receptor. These are either a CD4 or CD8 molecule:
- CD4 is present on T helper cells and only binds to antigen-MHC II complexes.
- CD8 is present on cytotoxic T cells and only binds to antigen-MHC I complexes.
This, therefore, leads to very different effects. Antigens presented with MHC II will activate T helper cells and antigens presented with MHC I activate cytotoxic T cells. Cytotoxic T cells will kill the cells that they recognise, whereas T helper cells have a broader range of effects on the presenting cell such as activation to produce antibodies (in the case of B cells) or activation of macrophages to kill their intracellular pathogens.
Clinical Relevance – Autoimmune disease
It is important to note that APCs may deliver foreign antigens or self-antigens. In the case of autoimmune diseases, self-antigens are presented to T cells, which then initiates an immune response against our own tissues.
For example, in Graves’ disease , TSHR (thyroid stimulating hormone receptor) acts as a self-antigen and is presented to T cells. This then activates B cells to produce autoantibodies against TSHRs in the thyroid. This results in the activation of TSHRs leading to hyperthyroidism and a possible goitre.
[start-clinical]
Clinical Relevance - Autoimmune disease
[end-clinical]
Found an error? Is our article missing some key information? Make the changes yourself here!
Once you've finished editing, click 'Submit for Review', and your changes will be reviewed by our team before publishing on the site.
We use cookies to improve your experience on our site and to show you relevant advertising. To find out more, read our privacy policy .
Privacy Overview
BiteSized Immunology: Systems & Processes

Antigen Processing and Presentation
In order to be capable of engaging the key elements of adaptive immunity (specificity, memory, diversity, self/nonself discrimination), antigens have to be processed and presented to immune cells. Antigen presentation is mediated by MHC class I molecules , and the class II molecules found on the surface of antigen-presenting cells (APCs) and certain other cells.
MHC class I and class II molecules are similar in function: they deliver short peptides to the cell surface allowing these peptides to be recognised by CD8+ (cytotoxic) and CD4+ (helper) T cells, respectively. The difference is that the peptides originate from different sources – endogenous, or intracellular , for MHC class I; and exogenous, or extracellular for MHC class II. There is also so called cross-presentation in which exogenous antigens can be presented by MHC class I molecules. Endogenous antigens can also be presented by MHC class II when they are degraded through autophagy.

MHC class I presentation
MHC class I molecules are expressed by all nucleated cells. MHC class I molecules are assembled in the endoplasmic reticulum (ER) and consist of two types of chain – a polymorphic heavy chain and a chain called β2-microglobulin. The heavy chain is stabilised by the chaperone calnexin , prior to association with the β2-microglobulin. Without peptides, these molecules are stabilised by chaperone proteins : calreticulin, Erp57, protein disulfide isomerase (PDI) and tapasin. The complex of TAP, tapasin, MHC class I, ERp57 and calreticulin is called the peptide-loading complex (PLC). Tapasin interacts with the transport protein TAP (transporter associated with antigen presentation) which translocates peptides from the cytoplasm into the ER. Prior to entering the ER, peptides are derived from the degradation of proteins, which can be of viral- or self origin. Degradation of proteins is mediated by cytosolic- and nuclear proteasomes, and the resulting peptides are translocated into the ER by means of TAP. TAP translocates peptides of 8 –16 amino acids and they may require additional trimming in the ER before binding to MHC class I molecules. This is possibly due to the presence of ER aminopeptidase (ERAAP) associated with antigen processing.
It should be noted that 30–70% of proteins are immediately degraded after synthesis (they are called DRiPs – defective ribosomal products, and they are the result of defective transcription or translation). This process allows viral peptides to be presented very quickly – for example, influenza virus can be recognised by T cells approximately 1.5 hours post-infection. When peptides bind to MHC class I molecules, the chaperones are released and peptide–MHC class I complexes leave the ER for presentation at the cell surface. In some cases, peptides fail to associate with MHC class I and they have to be returned to the cytosol for degradation. Some MHC class I molecules never bind peptides and they are also degraded by the ER-associated protein degradation (ERAD) system.
There are different proteasomes that generate peptides for MHC class-I presentation: 26S proteasome , which is expressed by most cells; the immunoproteasome, which is expressed by many immune cells; and the thymic-specific proteasome expressed by thymic epithelial cells.
Antigen presentation
On the surface of a single cell, MHC class I molecules provide a readout of the expression level of up to 10,000 proteins. This array is interpreted by cytotoxic T lymphocytes and Natural Killer cells, allowing them to monitor the events inside the cell and detect infection and tumorigenesis.
MHC class I complexes at the cell surface may dissociate as time passes and the heavy chain can be internalised. When MHC class I molecules are internalised into the endosome, they enter the MHC class-II presentation pathway. Some of the MHC class I molecules can be recycled and present endosomal peptides as a part of a process which is called cross-presentation .
The usual process of antigen presentation through the MHC I molecule is based on an interaction between the T-cell receptor and a peptide bound to the MHC class I molecule. There is also an interaction between the CD8+ molecule on the surface of the T cell and non-peptide binding regions on the MHC class I molecule. Thus, peptide presented in complex with MHC class I can only be recognised by CD8+ T cells. This interaction is a part of so-called ‘three-signal activation model’, and actually represents the first signal. The next signal is the interaction between CD80/86 on the APC and CD28 on the surface of the T cell, followed by a third signal – the production of cytokines by the APC which fully activates the T cell to provide a specific response.
MHC class I polymorphism
Human MHC class I molecules are encoded by a series of genes – HLA-A, HLA-B and HLA-C (HLA stands for ‘Human Leukocyte Antigen’, which is the human equivalent of MHC molecules found in most vertebrates). These genes are highly polymorphic, which means that each individual has his/her own HLA allele set. The consequences of these polymorphisms are differential susceptibilities to infection and autoimmune diseases that may result from the high diversity of peptides that can bind to MHC class I in different individuals. Also, MHC class I polymorphisms make it virtually impossible to have a perfect tissue match between donor and recipient, and thus are responsible for graft rejection.

MHC class II presentation
MHC class II molecules are expressed by APCs, such as dendritic cells (DC), macrophages and B cells (and, under IFNγ stimuli, by mesenchymal stromal cells, fibroblasts and endothelial cells, as well as by epithelial cells and enteric glial cells). MHC class II molecules bind to peptides that are derived from proteins degraded in the endocytic pathway. MHC class II complexes consists of α- and β-chains that are assembled in the ER and are stabilised by invariant chain (Ii). The complex of MHC class II and Ii is transported through the Golgi into a compartment which is termed the MHC class II compartment (MIIC). Due to acidic pH, proteases cathepsin S and cathepsin L are activated and digest Ii, leaving a residual class II-associated Ii peptide (CLIP) in the peptide-binding groove of the MHC class II. Later, the CLIP is exchanged for an antigenic peptide derived from a protein degraded in the endosomal pathway. This process requires the chaperone HLA-DM, and, in the case of B cells, the HLA-DO molecule. MHC class II molecules loaded with foreign peptide are then transported to the cell membrane to present their cargo to CD4+ T cells. Thereafter, the process of antigen presentation by means of MHC class II molecules basically follows the same pattern as for MHC class I presentation.
As opposed to MHC class I, MHC class II molecules do not dissociate at the plasma membrane. The mechanisms that control MHC class II degradation have not been established yet, but MHC class II molecules can be ubiquitinised and then internalised in an endocytic pathway.
MHC class II polymorphism
Like the MHC class I heavy chain, human MHC class II molecules are encoded by three polymorphic genes: HLA-DR, HLA-DQ and HLA-DP. Different MHC class II alleles can be used as genetic markers for several autoimmune diseases, possibly owing to the peptides that they present.
Module 20: The Immune System
Antigen-presenting cells, learning outcomes.
- Describe the structure and function of antigen-presenting cells
Unlike NK cells of the innate immune system, B cells (B lymphocytes) are a type of white blood cell that gives rise to antibodies, whereas T cells (T lymphocytes) are a type of white blood cell that plays an important role in the immune response. T cells are a key component in the cell-mediated response—the specific immune response that utilizes T cells to neutralize cells that have been infected with viruses and certain bacteria. There are three types of T cells: cytotoxic, helper, and suppressor T cells. Cytotoxic T cells destroy virus-infected cells in the cell-mediated immune response, and helper T cells play a part in activating both the antibody and the cell-mediated immune responses. Suppressor T cells deactivate T cells and B cells when needed, and thus prevent the immune response from becoming too intense.
An antigen is a foreign or “non-self” macromolecule that reacts with cells of the immune system. Not all antigens will provoke a response. For instance, individuals produce innumerable “self” antigens and are constantly exposed to harmless foreign antigens, such as food proteins, pollen, or dust components. The suppression of immune responses to harmless macromolecules is highly regulated and typically prevents processes that could be damaging to the host, known as tolerance.
The innate immune system contains cells that detect potentially harmful antigens, and then inform the adaptive immune response about the presence of these antigens. An antigen-presenting cell (APC) is an immune cell that detects, engulfs, and informs the adaptive immune response about an infection. When a pathogen is detected, these APCs will phagocytose the pathogen and digest it to form many different fragments of the antigen. Antigen fragments will then be transported to the surface of the APC, where they will serve as an indicator to other immune cells. Dendritic cells are immune cells that process antigen material; they are present in the skin (Langerhans cells) and the lining of the nose, lungs, stomach, and intestines. Sometimes a dendritic cell presents on the surface of other cells to induce an immune response, thus functioning as an antigen-presenting cell. Macrophages also function as APCs. Before activation and differentiation, B cells can also function as APCs.
After phagocytosis by APCs, the phagocytic vesicle fuses with an intracellular lysosome forming phagolysosome. Within the phagolysosome, the components are broken down into fragments; the fragments are then loaded onto MHC class I or MHC class II molecules and are transported to the cell surface for antigen presentation, as illustrated in Figure 1. Note that T lymphocytes cannot properly respond to the antigen unless it is processed and embedded in an MHC II molecule. APCs express MHC on their surfaces, and when combined with a foreign antigen, these complexes signal a “non-self” invader. Once the fragment of antigen is embedded in the MHC II molecule, the immune cell can respond. Helper T- cells are one of the main lymphocytes that respond to antigen-presenting cells. Recall that all other nucleated cells of the body expressed MHC I molecules, which signal “healthy” or “normal.”

Figure 1. An APC, such as a macrophage, engulfs and digests a foreign bacterium. An antigen from the bacterium is presented on the cell surface in conjunction with an MHC II molecule Lymphocytes of the adaptive immune response interact with antigen-embedded MHC II molecules to mature into functional immune cells.
Candela Citations
- Biology 2e. Provided by : OpenStax. Located at : http://cnx.org/contents/[email protected] . License : CC BY: Attribution . License Terms : Access for free at https://openstax.org/books/biology-2e/pages/1-introduction


- school Campus Bookshelves
- menu_book Bookshelves
- perm_media Learning Objects
- login Login
- how_to_reg Request Instructor Account
- hub Instructor Commons
Margin Size
- Download Page (PDF)
- Download Full Book (PDF)
- Periodic Table
- Physics Constants
- Scientific Calculator
- Reference & Cite
- Tools expand_more
- Readability
selected template will load here
This action is not available.
20.3E: Antigen-Presenting Cells
- Last updated
- Save as PDF
- Page ID 7949
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
Antigen presentation is a process by which immune cells capture antigens and then enable their recognition by T cells.
Learning Objectives
- Describe the role of antigen-presenting cells
- The host’s cells express “self” antigens that identify them as such. These antigens are different from those in bacteria (“non-self” antigens) and in virus-infected host cells (“missing-self”).
- Antigen presentation consists of pathogen recognition, phagocytosis of the pathogen or its molecular components, processing of the antigen, and then presentation of the antigen to naive T cells.
- The T cell receptor is restricted to recognizing antigenic peptides only when bound to appropriate molecules of the major histocompatibility complex (MHC), also known in humans as human leukocyte antigen (HLA).
- Helper T cells recieve antigens from MHC II on an APC, while cytotoxic T cells recieve antigens from MHC I. Helper T cells present their antigen to B cells as well.Dendritic cells, B cells, and macrophages play a major role in the innate response, and are the primary antigen-presenting cells (APC).
- APCs use toll-like receptors to identify PAMPS and DAMPs, which are signs of an infection and may be processed into antigen peptides if phagocytized. Most APCs cannot tell the difference between different types of antigens like B and T cells can.
- damage-associated molecular pattern : Protein or nucleic acid based signs of pathogen induced damage. Protein DAMPs may be phagocytized and processed for antigen presentation.
- cytotoxic : A population of T cells specialized for inducing the deaths of other cells.
Antigen presentation is a process in the body’s immune system by which macrophages, dendritic cells and other cell types capture antigens, then present them to naive T-cells. The basis of adaptive immunity lies in the capacity of immune cells to distinguish between the body’s own cells and infectious pathogens. The host’s cells express “self” antigens that identify them as belonging to the self. These antigens are different from those in bacteria (“non-self” antigens) or in virally-infected host cells (“missing-self”). Antigen presentation broadly consists of pathogen recognition, phagocytosis of the pathogen or its molecular components, processing of the antigen, and then presentation of the antigen to naive (mature but not yet activated) T cells. The ability of the adaptive immune system to fight off pathogens and end an infection depends on antigen presentation.
Antigen Presenting Cells
Antigen Presenting Cells (APCs) are cells that capture antigens from within the body, and present them to naive T-cells. Many immune system cells can present antigens, but the most common types are macrophages and dendritic cells, which are two types of terminally differentiated leukocytes that arise from monocytes. Both of these APCs perform many immune functions that are important for both innate and adaptive immunity, such as removing leftover pathogens and dead neutrophils after an inflammatory response. Dendritic cells (DCs) are generally found in tissues that have contact with the external environment (such as the skin or respiratory epithelium) while macrophages are found in almost all tissues. Some types of B cells may also present antigens as well, though it is not their primary function.
APCs phagocytize exogenous pathogens such as bacteria, parasites, and toxins in the tissues and then migrate, via chemokine signals, to lymph nodes that contain naive T cells. During migration, APCs undergo a process of maturation in which they digest phagocytized pathogens and begin to express the antigen in the form of a peptide on their MHC complexes, which enables them to present the antigen to naive T cells. The antigen digestion phase is also called “antigen processing,” because it prepares the antigens for presentation. This MHC:antigen complex is then recognized by T cells passing through the lymph node. Exogenous antigens are usually displayed on MHC Class II molecules, which interact with CD4+ helper T cells.
This maturation process is dependent on signaling from other pathogen-associated molecular pattern (PAMP) molecules (such as a toxin or component of a cell membrane from a pathogen) through pattern recognition receptors (PRRs), which are received by Toll-like receptors on the DC’s body. They may also recognize damage-associated molecular pattern (DAMP) molecules, which include degraded proteins or nucleic acids released from cells that undergo necrosis. PAMPs and DAMPS are not technically considered antigens themselves, but instead are signs of pathogen presence that alert APCs through Toll-like receptor binding. However if a DC phagocytzes a PAMP or DAMP, it could be used as an antigen during antigen presentation. APCs are unable to distinguish between different types of antigens themselves, but B and T cells can due to their specificity.
Antigen Presentation
T cells must be presented with antigens in order to perform immune system functions. The T cell receptor is restricted to recognizing antigenic peptides only when bound to appropriate molecules of the MHC complexes on APCs, also known in humans as Human leukocyte antigen (HLA).
Several different types of T cell can be activated by APCs, and each type of T cell is specially equipped to deal with different pathogens, whether the pathogen is bacterial, viral or a toxin. The type of T cell activated, and therefore the type of response generated, depends on which MHC complex the processed antigen-peptide binds to.
MHC Class I molecules present antigen to CD8+ cytotoxic T cells, while MHC class II molecules present antigen to CD4+ helper T cells. With the exception of some cell types (such as erythrocytes), Class I MHC is expressed by almost all host cells. Cytotoxic T cells (also known as TC, killer T cell, or cytotoxic T-lymphocyte (CTL)) are a population of T cells that are specialized for inducing the death of other cells. Recognition of antigenic peptides through Class I by CTLs leads to the killing of the target cell, which is infected by virus, intracytoplasmic bacterium, or are otherwise damaged or dysfunctional. Additionally, some helper T cells will present their antigen to B cells, which will activate their proliferation response.
Antigen presentation : In the upper pathway; foreign protein or antigen (1) is taken up by an antigen-presenting cell (2). The antigen is processed and displayed on a MHC II molecule (3), which interacts with a T helper cell (4). In the lower pathway; whole foreign proteins are bound by membrane antibodies (5) and presented to B lymphocytes (6), which process (7) and present antigen on MHC II (8) to a previously activated T helper cell (10), spurring the production of antigen-specific antibodies (9).
LICENSES AND ATTRIBUTIONS
CC LICENSED CONTENT, SHARED PREVIOUSLY
- Curation and Revision. Authored by : Boundless.com. Provided by : Boundless.com. License : CC BY-SA: Attribution-ShareAlike
CC LICENSED CONTENT, SPECIFIC ATTRIBUTION
- antigen. Provided by : Wiktionary. Located at : en.wiktionary.org/wiki/antigen . License : CC BY-SA: Attribution-ShareAlike
- Adaptive immune system. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Adaptive_immune_system . License : CC BY-SA: Attribution-ShareAlike
- antibody. Provided by : Wiktionary. Located at : en.wiktionary.org/wiki/antibody . License : CC BY-SA: Attribution-ShareAlike
- macrophage. Provided by : Wiktionary. Located at : en.wiktionary.org/wiki/macrophage . License : CC BY-SA: Attribution-ShareAlike
- Antibody. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/File:Antibody.jpg . License : Public Domain: No Known Copyright
- Antigen presentation. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/File:Antigen_presentation.svg . License : Public Domain: No Known Copyright
- Immune system. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Immune_system%23Innate_immune_system . License : CC BY-SA: Attribution-ShareAlike
- Lymphocyte_activation_simple.png. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Immune_system#/media/File:Lymphocyte_activation_simple.png . License : CC BY-SA: Attribution-ShareAlike
- T cell. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/T_cell . License : CC BY-SA: Attribution-ShareAlike
- B cell. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/B_cell . License : CC BY-SA: Attribution-ShareAlike
- Immune cells. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Immune_cells . License : CC BY-SA: Attribution-ShareAlike
- thymus. Provided by : Wiktionary. Located at : en.wiktionary.org/wiki/thymus . License : CC BY-SA: Attribution-ShareAlike
- Red White Blood cells. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/File:Red_White_Blood_cells.jpg . License : Public Domain: No Known Copyright
- Lymphocyte. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Lymphocyte . License : CC BY-SA: Attribution-ShareAlike
- B cells. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/B%20cells . License : CC BY-SA: Attribution-ShareAlike
- natural killer (NK) cells. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/natural...20(NK)%20cells . License : CC BY-SA: Attribution-ShareAlike
- T cells. Provided by : Wiktionary. Located at : en.wiktionary.org/wiki/T_cells . License : CC BY-SA: Attribution-ShareAlike
- Lymphocyte. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Lymphocyte . License : Public Domain: No Known Copyright
- Antigen presentation. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Antigen_presentation . License : CC BY-SA: Attribution-ShareAlike
- cytotoxic. Provided by : Wiktionary. Located at : en.wiktionary.org/wiki/cytotoxic . License : CC BY-SA: Attribution-ShareAlike
- Antigen presentation. Provided by : Wikipedia. Located at : en.Wikipedia.org/wiki/Antigen_presentation . License : Public Domain: No Known Copyright

- school Campus Bookshelves
- menu_book Bookshelves
- perm_media Learning Objects
- login Login
- how_to_reg Request Instructor Account
- hub Instructor Commons
Margin Size
- Download Page (PDF)
- Download Full Book (PDF)
- Periodic Table
- Physics Constants
- Scientific Calculator
- Reference & Cite
- Tools expand_more
- Readability
selected template will load here
This action is not available.
15.4M: Antigen Presentation
- Last updated
- Save as PDF
- Page ID 5449

- John W. Kimball
- Tufts University & Harvard
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
Antigens are macromolecules that elicit an immune response in the body. Antigens can be proteins, polysaccharides, conjugates of lipids with proteins (lipoproteins) and polysaccharides (glycolipids). Most of this page will describe how protein antigens are presented to the immune system. The presentation of lipid and polysaccharide antigens will be mentioned at the end. It will be helpful to distinguish between two limiting cases.
Antigens that enter the body from the environment; these would include inhaled macromolecules (e.g., proteins on cat hairs that can trigger an attack of asthma in susceptible people), ingested macromolecules (e.g., shellfish proteins that trigger an allergic response in susceptible people), and molecules that are introduced beneath the skin (e.g., on a splinter or in an injected vaccine). Alternatively, antigens can be generated within the cells of the body; these would include proteins encoded by the genes of viruses that have infected a cell and aberrant proteins that are encoded by mutant genes; such as mutated genes in cancer cells. In all cases, however, the initial immune response to any antigen absolutely requires that the antigen be recognized by a T lymphocyte ("T cell"). The truth of this rule is clearly demonstrated in AIDS : the infections (viral or fungal or bacterial) that so often claim the life of AIDS patients do so when the patient has lost virtually all of his or her CD4 + T cells. The two categories of antigens are processed and presented to T cells by quite different mechanisms.
Exogenous Antigens
Exogenous antigens (inhaled, ingested, or injected) are taken up by antigen-presenting cells (APCs). These include phagocytic cells like dendritic cells and macrophages and B lymphocytes ("B cells") which are responsible for producing antibodies against the antigen. Antigen-presenting cells
- engulf the antigen by endocytosis
- endosome fuses with a lysosome where the antigen is degraded into fragments (e.g. short peptides)
- these antigenic peptides are then displayed at the surface of the cell nestled within a class II histocompatibility molecule .
- they may be recognized by CD4 + T cells
The Class I Pathway
Class I histocompatibility molecules are transmembrane proteins expressed at the cell surface. Like all transmembrane proteins, they are synthesized by ribosomes on the rough endoplasmic reticulum (RER) and assembled within its lumen. There are three subunits in each class I histocompatibility molecule:
- the transmembrane polypeptide (called the "heavy chain")
- the antigenic peptide
- beta-2 microglobulin
All of these must be present within the lumen of the endoplasmic reticulum if they are to assemble correctly and move through the Golgi apparatus to the cell surface. The Problem: proteins encoded by the genes of an infecting virus are synthesized in the cytosol . How to get them into the endoplasmic reticulum?
The Solution: TAP (= t ransporter a ssociated with antigen p rocessing).
- Viral proteins in the cytosol are degraded by proteasomes into viral peptides.
- The peptides are picked up by TAP proteins embedded in the membrane of the endoplasmic reticulum.
- Using the energy of ATP, the peptides are pumped into the lumen of the endoplasmic reticulum where they assemble with the transmembrane polypeptide and beta-2 microglobulin.
- This trimolecular complex then moves through the Golgi apparatus and is inserted in the plasma membrane.
- The complex can be bound by a T cell with a receptor ( TCR ) able to bind the peptide and flanking portions of the histocompatibility molecule (the hot dog in the bun) and CD8 molecules that bind the CD8 receptor (shown above as a gray hemisphere) on the histocompatibility molecule.
The Class II Pathway
Class II histocompatibility molecules consist of two transmembrane polypeptides and a third molecule nestled in the groove they form. All three components of this complex must be present in the endoplasmic reticulum for proper assembly. But antigenic peptides are not transported to the endoplasmic reticulum, so a protein called the invariant chain (" Ii ") temporarily occupies the groove.
- The two chains of the class II molecule are inserted into the membrane of the endoplasmic reticulum.
- They bind (in their groove) one molecule of invariant chain.
- This trimolecular complex is transported through the Golgi apparatus and into vesicles called lysosomes.
Meanwhile foreign antigenic material is engulfed by endocytosis forming endosomes . These also fuse with lysosomes. Then,
- The antigen is digested into fragments.
- The invariant (Ii) chain is digested.
- This frees the groove for occupancy by the antigenic fragment.
- The vesicles move to the plasma membrane and the complex is displayed at the cell surface.
- a receptor ( TCR ) able to bind the peptide and flanking portions of the histocompatibility molecule (the hot dog in the bun) and
- CD4 molecules that bind the CD4 receptor (shown above as a yellow triangle) found on all class II histocompatibility molecules.
Interconnections Between the Class I and Class II Pathways
Cross-presentation: transferring exogenous antigens to the class i pathway.
Cross-presentation is the transferring of extracellular antigens like bacteria, some tumor antigens, and antigens in cells infected by viruses into the class I pathway for stimulation of CD8 + cytotoxic T cells (CTL). Only certain "professional" antigen-presenting cells (APCs) like dendritic cells can do this - use the class I as well as the class II pathways of antigen presentation.
Cross-presentation following infection by viruses is important because:
- Most viruses infect cells other than APCs (e.g., liver cells, epithelial cells of the lung) (and, of course, are intracellular in these).
- While viral antigens displayed on the surface of any infected cell can serve as targets for cytotoxic T cells (CTLs),
- the lack of any costimulatory molecules on the cell surface makes them poor stimulants for the development of clones of CTLs in the first place.
However, when an infected cell dies, it can be engulfed by a professional APC, and the antigens within it can enter the class I pathway. One mechanism:
- The dead cell is engulfed by endocytosis.
- The endosome that forms fuses with a lysosome and degradation of the dead cell begins.
- Antigens pass into the cytosol and are degraded in proteasomes.
- The peptides formed are then are picked up by TAP and inserted into class I MHC molecules and displayed at the cell surface — along with the costimulatory molecules needed to start a vigorous clonal expansion of CD8 + cytotoxic T cells.
Diverting Antigens from the Class I to the Class II Pathway
Autophagy provides a mechanism by which cells can transfer endogenous (intracellular) antigens into the class II pathway, for example
- self-proteins so as to be able to delete CD4 + T cells with receptors capable of attacking them and thus potentially capable of causing autoimmunity
- proteins synthesized by an infecting virus. In this way viral infection can generate CD4 + T cells as well as cytotoxic T cells (CD8 + )
B Lymphocytes: A Special Case
B lymphocytes are both antigen-receiving and antigen-presenting cells. They bind intact antigens (e.g., virus particles, proteins) with their B cell receptor (BCR). They can come in contact with these antigens by encountering them in the surrounding lymph or by being presented them by macrophages or dendritic cells. B lymphocytes process antigen by the class II pathway for presentation to T cells.
The process:
- B cells engulf antigen by receptor-mediated endocytosis
- The B cell receptors for antigen ( BCR s) are antibodies anchored in the plasma membrane.
- The affinity of these for an epitope on an antigen may be so high that the B cell can bind and internalize the antigen when it is present in body fluids in concentrations thousands of times smaller than a macrophage would need.
- The remaining steps of antigen processing occur by the same class II pathway described above for macrophages producing
- fragments of antigen displayed at the cell surface nestled in the groove of class II histocompatibility molecules.
- A CD4 + T cell that recognizes the displayed antigen is stimulated to release lymphokines.
- These, in turn, stimulate the B cell to enter the cell cycle.
- Because of the part they play in stimulating B cells, these CD4 + T cells are called Helper T cells (" Th ").
- The B cell grows into a clone of cells (called plasma cells )
- These synthesize receptors ( BCR s) with the identical binding site for the epitope but without the transmembrane tail.
- These antibodies are secreted into the surroundings.
Lipid and Polysaccharide Antigens
Lipid antigens.
- Lipid antigens are presented to T cells by cell-surface molecules designated CD1 ("cluster of differentiation" 1).
- Antigen-presenting cells express several different forms of CD1 at their surface. Each is probably specialized to bind a particular type of lipid antigen (e.g. lipopeptide vs glycolipid).
- The exposed surface of CD1 molecules forms an antigen-binding groove much like that of MHC molecules except that
- the amino acids in the groove are more hydrophobic than those in MHC molecules.
- Like protein antigens, lipid antigens are also presented as fragments, i.e., as a "hot dog in a bun".
Polysaccharide Antigens
Some bacterial polysaccharides ingested by APCs
- can be degraded in their lysosomes
- and presented to T cells by MHC class II molecules.
The binding of a T cell to an antigen-presenting cell (APC) is by itself not enough to activate the T cell and turn it into an effector cell: one able to, for examples,
- kill the APC (CD8 + cytotoxic T lymphocytes [CTLs])
- carry out cell-mediated immune reactions (CD4 + Th1 cells)
- provide help to B cells (CD4 + Th2 cells)
In order to become activated, the T cell must not only bind to the epitope (MHC-peptide) with its TCR but also receive a second signal from the APC. The receipt of this second signal is called costimulation . Among the most important of these costimulators are molecules on the APC designated B7 and their ligand on the T cell designated CD28 . The binding of CD28 to B7 provides the second signal needed to activate the T cell.
Although T cells may encounter self antigens in body tissues, they will not respond unless they receive a second signal. In fact, binding of their TCR ("signal one") without "signal two" causes them to self-destruct by apoptosis. Most of the time, the cells presenting the body's own antigens either
- fail to provide signal two or
- transmit an as-yet-unidentified second signal that turns the T cell into a regulatory T cell ( Treg ) that suppresses immune responses.
In either case, self-tolerance results.
The Immune System Explainer
Immuno Studies World
Antigen Presenting Cells (APCs) and Their Role in Activation of The T cells Simplified

Please Share Content with Others: Sharing is Caring!
Antigen presenting cells (APCs) are important cells in the activation of T cells. That means that the cell-mediated branch of the adaptive immunity cannot be activated without APCs. The APCs are specialized immune cells that present antigens to T-cells.
These cells are involved in the activation of both innate and adaptive immunity . The link the two branches of the immune system. Therefore, APCs play a role in the regulation of the immune system by activating T-cells.
In this article, we shall discuss the different types of APCs, and how they process and present antigens to T cells. We shall also discuss how the T lymphocytes respond to the signals by APCs eventually becoming activated.
Types of Antigen Presenting Cells
Nearly all nucleated cells in your body can present antigens and hence serve as APCs. This is particularly the case with all virally infected cells and cancer cells . They can process and present antigen to CD8+ cytotoxic T lymphocytes.
To underscore the importance of APCs here, I need to mention that scientists have made artificial APCs that activate this type of T cells in the treatment of cancer.
Apart from this type of APCs, there are three types of professional APCs in our bodies that present antigens to the CD4+ T helper cells for recognition and subsequent amplification of the immune response.
The three include macrophages, dendritic cells, and the B cells. The first two are descendants of monocytes. The B cells emanate from the common lymphoid progenitor become plasma cells which produce antibodies.
The three types of APCs are termed professional because they can cross-present antigens through MHC class 1 pathway. Thats means they can present antigens in the context of both MHC class I and class II.
Exogenous antigens are presented to CD4+ T cells using the MHC class II pathway while the endogenous antigens are presented to the CD8+ cytotoxic T cells using MHC class I pathway.
Let’s now provide a little bit of details about the three types of APCs here:
Dendritic cells
Dendritic cells (DCs) as already mentioned develop from monocytes. That is to say that DCs are descendants of the common myeloid progenitor cells. The DCs are responsible for initiating immune responses by processing antigens and presenting them to T-cells.

When it comes to antigen presentation, DCs are the most efficient in cross-presentation . They can prime both the CD8+ cytotoxic T cells and the CD4+ T helper cells. They are professional APCs.
DCs have two major subtypes: myeloid dendritic cells (mDC) and plasmacytoid dendritic cells (pDC). The mDCs are found throughout the body and are specialized at capturing antigens and transporting them to lymph nodes where they present them to T-cells.
On the other, pDCs reside primarily in the spleen and bone marrow. The dendric cells phagocytose antigens, process them before they can present them to T cells through MHC molecules.
Macrophages
Macrophages are phagocytic cells that engulf foreign particles and pathogens. They are derived from monocytes and are classified based on their morphology and function.
There are three main types of macrophages : M0, M1, and M2. M0 macrophages are non-activated and do not express any specific markers.
M1 macrophages are activated by interferon gamma (IFNγ), lipopolysaccharide (LPS), and tumor necrosis factor alpha (TNFα).
M1 macrophages produce proinflammatory cytokines that include IL-12, IL-23, and TNFα. As for the M2 macrophages, they are activated via Th2 cytokines including IL4 and IL13.

M2 macrophage activation results in the production of anti-inflammatory cytokines such as TGFβ and IL10.
These cytokines are important in slowing down inflammation and hence avert the inflammatory response-related cellular damage in your body.
B cells are white blood cells that are responsible for producing antibodies that fight off foreign invaders. However, they too process and present antigens to T cells.
However, different from other APCs the B cells use receptor-mediated endocytosis to capture the antigen, process and present them to T cells. There are two types of B cells, namely B lymphocytes and plasma cells.
B lymphocytes are the primary producers of antibodies. They are responsible in fighting off bacteria and viruses. Plasma cells are the final stage of B cell development and are the only ones involved in antibody production.

Therefore, plasma cells are responsible for secreting antibodies into your blood bloodstream. Those antibodies are of course then involved in several types of effector mechanisms resulting in the elimination of antigens.
The antibodies also known as immunoglobulins are of five classes including IgA, IgD, IgE, IgG and IgM. They have different properties with some like IgD and IgM bound on the B cell membrane while the rest are soluble free serum proteins in your blood circulation.
Antigen Processing
The antigen presenting cells engulf antigens, then catalytically process them into short peptides ready for presentation to the T cells.
The T cells cannot directly recognize and bind to antigens like other immune cells would do. The APCs possess major histocompatibility complex (MHC) molecules on their surface which they use to present antigens to the T cells.
The major histocompatibility complex (MHC) is a group of genes located on chromosome 6. MHC genes encode proteins that help the immune system distinguish between self and non-self.
This is certainly important to ensure that your immune system doesn’t fight your own cells which would result in autoimmunity . There are two types of MHC molecules involved in antigen presentation to T cells. These are MHC class I and MHC class II molecules.
MHC class II molecules are proteins on the surface of certain types of cells (e.g., Dendritic cells and macrophages) that present peptides to the CD4+ helper T-cells.
Peptides are short chains of amino acids that are produced inside your APCs during an infection. Helper T-cells recognize peptide fragments bound to MHC class II molecules on the surface of antigen presenting cells.
On the other hand, a CD8+ T-cell recognizes antigens presented by the MHC class I molecules. As stated earlier there are many cells in your body that express MHC class I molecules but the most notable ones are the virally infected cells and the cancerous cells.
T cell Activation by APCs
T cells activation occurs when APCs present antigens them. The process of activation of the T cells by the APCs is a two-signal process. These two signals must be delivered to ensure complete activation and functionality of the T cells.

In the case where signal 2 has not been fully delivered, the T cells instead of being activated become anergic or die by apoptosis. Anergy is a state of unresponsiveness and that means that such T cells would not help in clearing the antigen that triggered its activation in the first place.
This signal involves the interaction between the T cell receptor (TCR) on the T cells and MHC molecules on the surface of the APCs. During this process your T cells are said to have recognized the antigen.
That means we can simply call signal 1, the recognition stage of the T cell activation process. This stage is critical because without it then specific clones of T cells with specific TCRs for the antigen would not be selected.
This signal involves co-stimulation where CD28 markers on the surface of the T cells interact with the B-7 molecules on the APCs to amplify the activation of T cells. The B7 molecules on APCs include B7-1 (CD80) and B7-2 (CD86).
The absence of this signal leads to the ‘abortion’ of the T cell activation process. The cells being activated will go into a state of unresponsiveness called anergy. Anergy can also be result of upregulation of CTLA-4 (CD152) instead of CD28 for co-stimulation.
There could also be involvement of CD40L (CD154) on Th2 instead of CD28 in signal 2. CD40L interacts with its counter receptor on the B cells when the B cells serve as antigen presenting cells.
The presence of the CD40L-CD40 interactions leads to the ability of B cells to class switch in antibody production. The absence of this signal causes the lack of this important T cell-dependent effector mechanisms on the part of B cells.
Antigen presenting cells are very important cells of the immune system. The T cells which are the most important cells of the adaptive immune system cannot recognize antigens unless presented by the APCs. The APCs possess either MHC class I or II through which they interact with the TCRs on the surface of T cells.
A second signal where the B-7 molecules on the APCs interact with CD28 on the T cells help complete the activation of T cells. Without signal 2, the T cells become anergic and hence not helpful in eliminating the microorganisms or other antigens.
4 thoughts on “ Antigen Presenting Cells (APCs) and Their Role in Activation of The T cells Simplified ”
Never better than it is, , , 👌👌👌
Thanks a lot, Mary and always welcome to learn more…
very helpful indeed, salute sir
Leave a Reply Cancel reply
You must be logged in to post a comment.
IMAGES
COMMENTS
Antigen presentation is a vital immune process that is essential for T cell immune response triggering. Because T cells recognize only fragmented antigens displayed on cell surfaces, antigen processing must occur before the antigen fragment can be recognized by a T-cell receptor.
Jul 17, 2023 · This process of antigen presentation allows T cells to “see” what proteins are present in the body and to form an adaptive immune response against them. In this article, we shall discuss antigen processing, presentation, and recognition by T cells.
In order to be capable of engaging the key elements of adaptive immunity (specificity, memory, diversity, self/nonself discrimination), antigens have to be processed and presented to immune cells.
An antigen-presenting cell (APC) or accessory cell is a cell that displays an antigen bound by major histocompatibility complex (MHC) proteins on its surface; this process is known as antigen presentation.
antigen-presenting cell (APC): an immune cell that detects, engulfs, and informs the adaptive immune response about an infection. Major histocompatibility complex (MHC): a group of genes that code for proteins found on the surfaces of cells that help the immune system recognize foreign substances.
An antigen-presenting cell (APC) is an immune cell that detects, engulfs, and informs the adaptive immune response about an infection. When a pathogen is detected, these APCs will phagocytose the pathogen and digest it to form many different fragments of the antigen.
Antigen Recognition: how does the adaptive immune system “see” antigen? • B cells: – Antibody on B cells or free antibody can recognize intact antigen (i.e. soluble antigens, and cell surface antigens). – Proteins, nucleic acids, polysaccharides, lipids, and small molecules are antigenic for B cells.
Antigen presentation is a process in the body’s immune system by which macrophages, dendritic cells and other cell types capture antigens, then present them to naive T-cells. The basis of adaptive immunity lies in the capacity of immune cells to distinguish between the body’s own cells and infectious pathogens.
May 15, 2022 · Antigens are macromolecules that elicit an immune response in the body. Antigens can be proteins, polysaccharides, conjugates of lipids with proteins (lipoproteins) and polysaccharides (glycolipids). Most of this page will describe how protein antigens are presented to the immune system.
Oct 10, 2022 · The APCs are specialized immune cells that present antigens to T-cells. These cells are involved in the activation of both innate and adaptive immunity. The link the two branches of the immune system. Therefore, APCs play a role in the regulation of the immune system by activating T-cells.